Реакции катаболизма пуриновых нуклеотидов
Содержание книги
- Полиоловый путь превращения глюкозы
- Анаэробный распад глюкозы (гликолиз). Этапы. Гликолитическая оксидоредукция. Регуляция. Физиологическое значение анаэробного распада глюкозы. Баланс энергии.
- Аэробное окисление глюкозы. Изложить анаэробную фазу.
- Изменение скорости реакций цтк и причины накопления кетоновых тел при некоторых состояниях
- Мобилизация гликогена (гликогенолиз)
- Схематичное расположение дефектных ферментов при различных гликогенозах
- Включение глицерина в синтез глюкозы
- Высшие жирные кислоты, структура, свойства, биологическая роль. Понятие о полиненасыщенных жирных кислотах.
- Гликолипиды - рецепторные молекулы
- Липиды пищи, их характеристика. Суточная потребность в липидах.
- Полный ферментативный гидролиз триацилглицерола
- Нарушения переваривания и всасывания пищевых жиров.
- Ресинтез триацилглицеринов в стенке кишечника.
- Формирование кетоновых тел. Причины и последствия кетоза.
- Удлинение цепи жирных кислот
- Реакции синтеза таг из фосфатидной кислоты
- Холестерин, строение, свойства, основные этапы синтеза и его регуляция.
- Пищевые продукты - источники белков. Нормы белка в питании. Биологическая ценность белков. Понятие об азотистом балансе.
- Переваривание белков в желудке, характеристика ферментов. Роль соляной кислоты.
- Изменение кислотности в желудке
- Переваривание белков и полипептидов в кишечнике. Характеристика протеиназ.
- Возможные пути превращений аминокислот
- Схема реакции трансаминирования
- Декарбоксилирование аминокислот. Гистамин, серотонин и другие биогенные амины.
- Появление аммиака в клетках непрерывно
- Реакции глюкозо-аланинового цикла (выделен рамкой).
- Взаимосвязь обмена серина, глицина, метионина и цистеина
- Синтез креатина и креатинина, креатинфосфат - дополнительный источник энергии мышечного сокращения.
- Патология белкового обмена. Белковое голодание. Причины и последствия.
- Врожденные нарушения обмена некоторых аминокислот (фенилкетонурия, алкаптонурия, цистиноз и цистинурия).
- Распад нуклеиновых кислот в тканях. Катаболизм пуриновых оснований.
- Реакции катаболизма пуриновых нуклеотидов
- Источники атомов пуринового кольца
- Биосинтез пиримидиновых нуклеотидов в тканях.
- Нарушения порфиринового обмена. Порфирии.
- Этапы метаболизма билирубина в организме
- Схема патогенеза механической желтухи
- Патологические формы гемоглобина
- Белки сыворотки крови. Отдельные представители. Биологические функции.
- Гамма-глобулины - защитные белки
- Единицы измерения активности ферментов
- Роль почек в регуляции водно-солевого обмена. Вазопрессин, альдостерон, ренин-ангиотензиновая система.
- Физико-химические свойства мочи в норме и ее химический состав.
- У мочи есть и другие особенности
- Патологические компоненты мочи. Клиническое значение анализа мочи.
- Эндогенная вода. Компартментализация жидкостей в организме. Значение воды в организме.
- Биологическая роль, регуляция уровня.
- Биологическая роль, регуляция уровня. Ионы кальция необходимы для передачи нервного импульса
- Регуляция синтеза и секреции. Активируют: Гипокальциемия повышает гидроксилирование витамина D по С1 в почках через
- Железо, его концентрация в сыворотке крови. Биологическая роль. Определение железа в сыворотке крови.
Конечным продуктом распада пуринов мочевая кислота является у человека, приматов, большинства млекопитающих и птиц. У остальных живых организмов мочевая кислота превращается в аллантоин.
Распад нуклеиновых кислот в тканях. Катаболизм пиримидиновых нуклеотидов.
Пиримидины распадаются до аммиака, углекислого газа и воды
Распад пиримидиновых нуклеотидов происходит параллельно, с использованием одинаковых реакций и ферментов. Можно отметить несколько специфичных ферментов:
1. Фермент 5'-нуклеотидаза отщепляет 5'-фосфатную группу от ЦМФ, УМФ и ТМФ.
2. Дезаминаза проводит окислительное дезаминирование цитидина.
3. Нуклеозид-фосфорилаза удаляет рибозу от уридина и тимидина.
4. Дигидроурацил-дегидрогеназа – восстановление урацила и тимина.
5. Дигидропиримидиназа осуществляет гидролитическое расщепление пиримидинового кольца.
6. Далее происходит гидролитическое отщепление аммиака и углекислого газа.
7. После окончательного разрушения пиримидинового кольца появившиеся β-аминокислоты направляются в реакции трансаминирования, после чего соответствующие кетокислоты изомеризуются и далее сгорают в ЦТК.

Реакции катаболизма пиримидиновых нуклеотидов
Биосинтез пуриновых нуклеотидов в тканях. Регуляция.
Синтез пуриновых оснований происходит во всех клетках организма, главным образом в печени. Исключение составляют эритроциты, полиморфноядерные лейкоциты, лимфоциты.
Условно все реакции синтеза можно разделить на 4 этапа:
Синтез 5'-фосфорибозиламина
Первая реакция синтеза пуринов заключается в активации углерода в положении С1 рибозо-5-фосфата, это достигается синтезом 5-фосфорибозил-1-дифосфата (ФРДФ). Фосфорибозил-дифосфат является тем якорем, на основе которого синтезируется сложный пуриновый цикл.
Вторая реакция – это перенос NH2-группы глутамина на активированный атом С1 с образованием 5'-фосфорибозиламина. Указанная NH2-группа фосфорибозиламина уже принадлежит будущему пуриновому кольцу и ее азот будет атомом номер 9.

Реакции синтеза 5'-фосфорибозиламина
Параллельно фофорибозилдифосфат используется при синтезе пиримидиновых нуклеотидов. Он реагирует с оротовой кислотой и рибозо-5-фосфат связывается с ней, образуя оротидилмонофосфат.
Синтез инозинмонофосфата
5-фосфорибозиламин вовлекается в девять реакций, и в результате образуется первый пуриновый нуклеотид – инозинмонофосфорная кислота (ИМФ). В этих реакциях источниками атомов пуринового кольца являются глицин, аспартат, еще одна молекула глутамина, углекислый газ и производные тетрагидрофолиевой кислоты (ТГФК). В целом на синтез пуринового кольца затрачивается энергия 6 молекул АТФ.
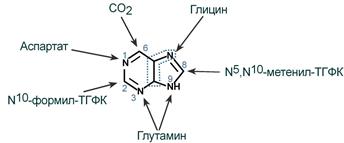
|