Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Общие молекулярные принципы действия наркотических веществСодержание книги
Поиск на нашем сайте Физиологические эффекты всех наркотических веществ реализуются на молекулярно-клеточном уровне, включающем взаимодействие с рецепторами различных типов, находящихся на поверхности нейронов ЦНС, а также развитием каскадных процессов внутриклеточной передачи сигналов, приводящих к физиологическому ответу клетки на уровне изменения активности и количества ферментов, а также регуляции экспрессии отдельных генов. Механизм действия и типы рецепторов, участвующих в реализации эффектов основных классов наркотических веществ приведены в Табл. 1. Табл. 1 Механизм действия некоторых наркотиков
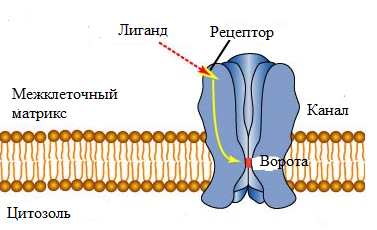
Gs – осуществляют сопряжение возбуждающих рецепторов с аденилатциклазой; Gi – осуществляют сопряжение тормозных рецепторов с аденилатциклазой; Go – осуществляют сопряжение рецепторов с ионными каналами; Gq – осуществляют сопряжение рецепторов, активирующих фосфолипазу C. Согласно существующей классификации, принято выделять два типа рецепторов: ионотропные и метаботропные. 1) Ионотропные рецепторы представляют собой крупные многосубъединичные белковые комплексы, состоящие из 4 - 5 индивидуальных белков, формирующих пронизывающий мембрану клетки ионный канал (Рис. 1).
Рис. 1 Строение ионотропного никотинового ацетилхолинового рецептора (nAChR). Ионотропный рецептор связвает лиганд, что приводит к открытию ионного канала посредством конформационных перестроек в структуре рецептора. рецептор состоит из пяти субъединиц, каждая из которых представлена четырьмя трансмембранными доменами. В отсутствии соответствующего лиганда, данный канал пребывает в закрытом состоянии, не позволяющем ионам проникать через мембрану клетки. Связывание лиганда с рецептором приводит к быстрым конформационным перестройкам в структуре рецептора, приводящим к открытию ионного канала и прохождению через него ионов по электрохимическому градиенту. Данные эффекты реализуются быстро, в течение миллисекунд, и исчезают, как только лиганд диссоциирует из комплекса с рецептором или же, когда происходит десенситизация рецептора по отношению к лиганду. Выделяют два семейства ионотропных рецепторов. Первое семейство включает рецептор ацетилхолина (ACh), никотиновый ацетилхолиновый рецептор (nAChR), рецептор γ-аминомасляной кислоты (ГАМК), рецептор ГАМКА, рецептор глицина, а также один из подклассов серотониновых рецепторов - 5НТ3. Второе семейство объединяет ионотропные рецепторы глутамата. В результате связывания нейромедиатора или наркотика с ионотропным рецептором происходит деполяризация мембраны и активация различных белков. Например, никотин связывается с рецептором nAChR, образующим Na+-канал. Бензодиазепины, барбитураты и этанол связываются с ГАМКА-рецепторами, содержащими Cl--каналы, в результате чего в клетку начинают проникать ионы Cl-. Этанол и фенциклидин ингибируют глутаматные рецепторы, чувствительные к N-метил-D-аспартату (NMDA-рецепторы), сопряженные с Ca2+-и Na+-каналами. 2) Метаботропные рецепторы, в отличие от ионотропных, обычно представлены единственным полипептидом, который передает сигнал внутрь клетки посредством связывания и активации ГТФ-связывающих белков (G-белков). Сигналы, передаваемые посредством метаботропных рецепторов реализуются более длительно (от нескольких десятков секунд до часов), вследствие осуществления большого количества последовательных ферментативных реакций, необходимых для осуществления эффекта. В настоящее время, данную группу рецепторов принято объединять в семейство GPCR (англ. G-protein coupled receptors). Данная группа рецепторов известна также под названием рецепторов семейства Т7, поскольку все рецепторы данного семейства имеют в составе своих молекул 7 трансмембранных доменов. Строение GPCR-рецепторов показано на Рис. 2.
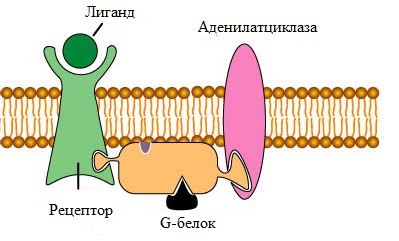
Рис. 2 Строение метаботропного рецептора. Метаботропный рецептор связывает лиганд. При этом в структуре рецептора происходят конформационные перестройки, приводящие к активации G -белка. Последние, в свою очередь, активируют ассоциированные с мембраной ферменты (например аденилатциклазу), генерирующие вторичные посредники в передаче сигнала внутрь клетки (например, цАМФ). К данному классу рецепторов принадлежат опиоидные рецепторы, β-адренергические рецепторы, дофаминовые рецепторы и канабиоидные рецепторы. Метаботропные рецепторы опосредуют медленные реакции. Ассоциированные с ними G-белки представляют собой гетеротримеры, состоящие из α-, β- и γ-субъединиц. α-субъединица обладает ГТФазной активностью. β- и γ-субъединицы, в отсутствие α-субъединицы способны образовывать βγ-димеры, не проявляющие собственной ферментативной активности, однако способные регулировать работу ионных каналов по принципу белок-белковых взаимодействий. В отсутствие лиганда, активный центр α-субъединицы содержит молекулу ГДФ и находится в комплексе с β- и γ-субъединицами. При связывании лиганда с рецептором, происходит активация ассоциированного с ним G-белка, что приводит к диссоциации α-субъединицы от βγ-димера. При этом происходит замещение молекулы ГДФ в активном центре α-субъединицы на молекулу ГТФ: α-ГДФ + ГТФ => α-ГТФ + ГДФ Как α-субъединица, так и βγ-димер могут активировать или ингибировать ферменты (аденилатциклазу или фосфолипазу С), в результате чего активируется (или подавляется) синтез вторичных посредников: цАМФ, инозитол-1,4,5-трифосфата (ИФ3) и диацилглицерола (ДАГ). Кроме того, β- и γ-субъединицы могут напрямую регулировать работу Ca2+, Na+ и K+-ионных каналов. Вторичные посредники также регулируют работу ионных каналов по механизму фосфорилирования/дефосфорилирования, посредством активации протеинкиназ. Активируемые вторичными посредниками протеинкиназы запускают каскадные процессы фосфорилирования различных белков в цитоплазме и ядре клетки, приводя к изменению активности ферментов и уровня экспрессии отдельных генов. Различные наркотические вещества реализуют свои эффекты через многочисленные рецепторы с использованием большого количества сигнальных механизмов. Связывание опиоидов с опиоидными рецепторами приводит к ингибированию аденилатциклазы и, соответственно, снижению уровня цАМФ и активности сАМФ-зависимой протеинкиназы А (ПКА/PKA). Каннабиноиды связываются с каннабиноидными рецепторами. Галлюциногены, такие как ЛСД, кетамин, скополамин и атропин, являются частичными агонистами серотониновых рецепторов. Амфетамины и кокаин активируют различные типы дофаминергических, адренергических и серотонинергических рецепторов и повышают уровень дофамина, норадреналина и серотонина в синапсах (Табл. 1).
|
|||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2021-05-12; просмотров: 186; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.20 (0.01 с.) |