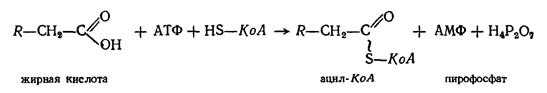Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Переваривание и всасывание липидов
Нейтральные жиры, поступающие в организм с пищей, подвергаются гидролитическому расщеплению на глицерин и жирные кислоты, катализируемому ферментами липазами.
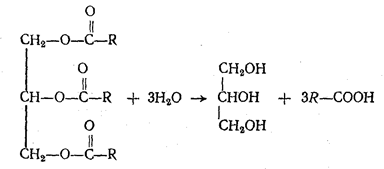
Подобно нейтральным жирам в кишечнике, расщепляются и фосфатиды. Гидролиз их, катализируемый фосфолипазами, приводит к образованию глицерина, жирных кислот, фосфорной кислоты и азотистого основания. Стериды, подвергаясь действию ферментов холестераз, гидролитически расщепляются на холестерин и соответствующую жирную кислоту. Решающее значение для переваривания и использования пищевых липидов имеет их эмульгирование, обеспечивающее большую доступность нерастворимых в воде липидов воздействию расщепляющих их липолитических ферментов, находящихся в водной фазе. Условий для эмульгирования в желудке нет. Поэтому липаза желудочного сока расщепляет лишь уже эмульгированные жиры пищи (например, жир молока). Основное переваривание жиров липазами сока поджелудочной железы и кишечника происходит в тонких кишках, чему способствуют соли желчных кислот, поступающие в кишечник с желчью и обладающие мощным эмульгирующим действием. Желчные кислоты по строению близки к холестерину. Это производные холановой кислоты (продукта окисления холестерина). Желчные кислоты располагаются на поверхности раздела между жиром и водой, причем в жировой фазе располагается гидрофобная (циклическая) часть молекулы, а в водной фазе — гидрофильный остаток. Это препятствует слиянию мелких капель в крупные. Кроме того, желчные кислоты вызывают уменьшение поверхностного натяжения на границе раздела фаз, способствуя неустойчивости крупных жировых капель и образованию из них более мелких, что увеличивает поверхность соприкосновения жировой и водной фаз. Увеличение поверхности жировых капель повышает эффективность действия липолитических ферментов. Благоприятные условия для их действия создаются и в ходе расщепления жиров, так как образующиеся при этом соли жирных кислот (мыла) также обладают эмульгирующими свойствами. Глицерин, образующийся при гидролизе липидов, хорошо растворяется в воде и легко всасывается слизистой оболочкой кишечника. Высшие жирные кислоты — стеариновая, пальмитиновая, олеиновая и др. в воде нерастворимы и могут всасываться лишь в виде водорастворимых комплексов с желчными кислотами так называемых холеиновых кислот.
Холеиновые кислоты, всасываясь, распадаются в эпителии кишечных ворсинок на свободные жирные и желчные кислоты. Часть освободившихся желчных кислот поступает обратно в просвет кишечника, способствуя дальнейшему всасыванию высших жирных кислот, другая их часть доставляется с кровью в печень, откуда переходит в состав желчи. Из всосавшихся жирных кислот и глицерина в кишечном эпителии происходит частичный ресинтез специфических триглицеридов; здесь же часть свободных жирных кислот включается в фосфатиды, преимущественно в лецитин, осуществляющий функцию транспорта жирных кислот. Большая часть жиров, ресинтезированных в эпителии кишечника или всосавшихся из полости кишечника без предварительного гидролиза благодаря высокой степени диспергирования, поступает в кровь через лимфатическую систему. Меньшая часть жиров (около 20—30%) поступает непосредственно в кровеносные капилляры и через систему воротной вены попадает в большой круг кровообращения. Основная масса жиров после всасывания откладывается в жировых депо организма: подкожной и забюшинной жировой клетчатке, сальнике. По мере надобности (например, при усиленном расходе энергии во время длительной мышечной деятельности, в условиях голодания и т. п.) происходит мобилизация запасных жиров из депо при участии липаз жировой ткани. Жиры и продукты липолиза — глицерин и жирные кислоты — поступают в кровь и ею разносятся к местам их использования. В плазме крови имеется сложная смесь липидов, в состав которой входят нейтральные жиры (триглицериды), фосфатиды, холестерин, его эфиры и свободные жирные кислоты. Транспорт нерастворимых в воде липидов осуществляется при участии белков плазмы и фосфатидов. Гидрофобные липиды с помощью фосфатидов (при посредстве их гидрофильных групп — фосфорной кислоты и азотистого основания) связаны с гидрофильным белком, обволакивающим (окутывающим) липиды. Образуются комплексные транспортные формы липидов — хиломикроны и липопротеиды плазмы крови. Диаметр первых — около 0,5 ммк. Вторые обладают меньшими размерами; их находят во фракциях глобулинов плазмы.
Свободные жирные кислоты транспортируются кровью в виде растворимых в воде комплексов с альбуминами плазмы.
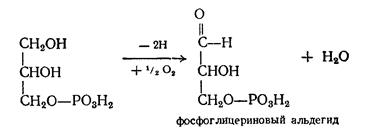
Окисление липидов в тканевом липолизе Продукты гидролиза липидов — жирные кислоты и глицерин — способны окисляться в тканях до СО2 и Н2О. Освобождающаяся при этом химическая энергия частично аккумулируется в богатых энергией фосфатных связях АТФ, частично рассеивается в виде тепла. Так как химическая природа жирных кислот и глицерина различна, различны и пути их окисления. Превращение глицерина Окисление глицерина начинается с его фосфорилирования; при взаимодействии глицерина с АТФ образуется фосфоглицерин.
Затем он дегидрогенируется, образуя фосфоглицериновый альдегид:
Фосфоглицериновый альдегид является одним из промежуточных продуктов гликолиза (см. превращения углеводов). Поэтому дальнейшие превращения протекают по схеме гликолиза, т.е. до лактата и пирувата, который превращается в ацетилКоА. Последний вступает в цикл трикарбоновых кислот, где ацетильный остаток окисляется до СО2 и Н2О. Превращение жирных кислот. В основе современных представлений о механизме окисления жирных кислот лежит теория, предложенная Ф.Кноопом в 1904 г. Им было экспериментально доказано, что молекулы жирной кислоты распадаются постепенно, укорачиваясь на 2 углеродных атома. Эта теория получила название теории В настоящее время установлено, что окислению жирных кислот в тканях предшествует их активация — взаимодействие с коферментом А в присутствии АТФ. Следствием этого является образование ацильного производного жирной кислоты, обладающего макроэргической связью:
Активированная жирная кислота (ацил-КоА) дегидрогенируется, вследствие чего возникает двойная связь между атомами углерода. Реакция катализируется ферментами, представляющими собою флавопротеиды (ФлП), содержащие в качестве простетической группы ФАД.
Затем по месту образования двойной связи, под влиянием фермента эноил-гидратазы, происходит присоединение молекулы воды (гидратация):
Образующий
Продукт этой реакции — кетоацилКоА расщепляется с одновременным присоединением новой молекулы КоА к укороченной на 2 атома углерода жирной кислоте:
В результате этой реакции образуются ацетилКоА и новый жирный ацилКоА, содержащий на два углеродных атома меньше, чем исходная жирная кислота. Он, в свою очередь, может подвергаться описанным выше превращениям, так что в конечном итоге вся углеродная цепь жирной кислоты распадается на двууглеродные фрагменты ацетилКоА. Образовавшийся при окислении жирных кислот ацетилКоА включается в цикл трикарбоновых кислот, где остаток уксусной кислоты окисляется до С02 и Н20, а КоА, освобождаясь, может вступать в конденсацию с новой молекулой жирной кислоты. При оксилении 1 молекулы ацетилКоА образуется 12 молекул АТФ. Кроме того, восстановленные ФАД и НАД, акцептирующие водород при дегидрогенированиях жирных кислот, окисляются в дыхательной цепи. При этом соответственно образуются 2 и 3 молекулы АТФ. В сумме полное окисление 1 двухуглеродного фрагмента любой жирной кислоты дает 17 молекул АТФ. Следовательно, при полном окислении 1 молекулы высокомолекулярной жирной кислоты, например пальмитиновой, образуется (8x17) 136 молекул АТФ. Из них 1 молекула расходуется на активацию жирной кислоты в самом начале процесса окисления.
|
||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-09-19; просмотров: 623; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.117.75.10 (0.011 с.) |

 -окисления, потому что окислению всегда подвергается второй от карбоксила углеродный атом (находящийся, следовательно, в
-окисления, потому что окислению всегда подвергается второй от карбоксила углеродный атом (находящийся, следовательно, в