
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Жизненный цикл клетки; пресинтетическая, синтетическая, постсинтетическая фаза; митоз, его характеристика.
Клеточный цикл – существование кл от деления до след деления или смерти. У одноклеточных особей клеточн цикл совпадает с жизнью особи. В непрерывно размножающихся кл клеточн цикл совпадает с митотическим циклом и состоит из 4х периодов (3 первых составляют интерфазу) со строгой последовательностью сменяющихся друг за другом: пресинтетический или постмитотический (G1), синтетический (S), постсинтетический или премитотическая (G2) и митоза (М). Различные периоды клеточн цикла отличаются др от др по общему содержанию в кл белка, ДНК, РНК и по уровню их синтеза. В G1 - периоде происходит активный рост и функционирование кл, обусловленные возобновлением транскрипции и накоплением синтезированных белков, а также подготовка к синтезу ДНК. Должна быть достигнута критическая масса цитоплазмы. В S - периоде происходит репликация ДНК. Продолжительность зав-т от скорости репликации ДНК (0,5 – 2 мкм/мин), от числа и величины репликонов, от числа включенных репликонов. Также идет удвоение материала хромосом. Для прохождения S – периода необходим синтез РНК и белков начавшийся еще в G1 -периоде. Параллельно синтезу ДНК идет интенсивно синтез гистонов в цитоплазме и происходит их миграция в ядро, где они связываются с ДНК. В S – периоде происходит синтез рРНК использующееся уже в G2 – периоде для синтеза белков, необходимых для митоза. G2 –период – осуществляется подготовка к делению, в том числе синтез белков веретена деления. Самая короткая из 3 стадий. Происходит синтез РНК и рРНК. В рез-те заключит этап клеточного цикла – митоза – редуплицированные хромосомы расходятся в дочерние кл. У покоящихся клеток выделяют G0–период – период покоя. Интерфаза период клеточного цикла м/у 2 последовательными митозами, по времени до 90% клеточн цикла. Митоз – непрямое деление. Биологическое значение состоит в строго одинаковом распределении редуплицированных хромосом м/у дочерними кл, что обеспечивает образование генетически равноценных клеток. Выделяют неск-ко фаз, постепенно и непрерывно переходящих др. в др: профаза, метафаза, анафаза, телофаза. Длительность стадий различна, зав-т от типа кл, физиологического состояния организма, но наиболее продолжительны первая и последняя. Профаза – конденсация хромосом, распад ядрышек и начало формирование веретена деления, снижение активности транскрипции (к концу профазы синтез РНК прекращается). Веретено деления образуется либо с участием центриолей, образуя митотический аппарат (в кл животных), либо без них (в кл высш раст). У простейших веретено деления может образовываться в ядре (каритотека во время митоза не разрушается) – закрытый митоз. Распад кариотеки на фрагменты, начин-ся беспорядочное движение хромосом в центральной части кл. Метафаза – завершается образование веретена деления. Хромосомы перестают двигаться и выстраиваются по экватору веретена, образуя экваториальную пластинку. Синтез белка снижен на 20 – 30 % по сравнению с интерфазой. В поздней метафазе хромосомы упорядочены (“застывшие”). Если посмотреть с полюса, то хромосомы лежат так, что центромерные участки расположены к центру веретена деления, плечи – к периферии – назыв-ся – “материнская звезда” (только у животных), у растений – по экватору без особого порядка. Завершается процессы обособления сестринских хроматид. Анафаза – самая короткая стадия. Хар-ся разделением сестринских хроматид и расхождением к противоположным полюсам кл. V=0,2 – 5 мкм/мин. Хромосомы приобретают V-образную форму. У некоторых высш раст нет выраженной центромеры, и волокно веретена контактирует со многими точками на поверхности хромосом. Иногда расхождение хромосом сопровождается расхождением полюсов. Телофаза – длится с момента прекращения движения хромосом до окончания процессов, связанных с реконструкцией дочерних ядер (деспирализации и активации хромосом, образование кариотеки, формирование ядрышек). Главное – разделение кл тела, цитотомия или цитокинез. У растений – образуется внутриклеточное образование клеточной перегородки, у животных – перетяжек, впячивания плазматической мембраны внутрь кл. Митоз не всегда заканчивается разделением кл. Так в эндосперме раст некоторое время может идти деление ядер без деления цитоплазмы. Также делятся ядра у плазмодиев миксомицетов.
4. Макромолекулы как основа организации биологических структур. Принципы образования вторичной, троичной и четвертичной структуры биомакромолекул и надмолекулярных структур.
Основу структурной организации живого составляют макромолекулы, прежде всего важнейшие биополимеры – белки и нуклеиновые кислоты. В макромолекулах происходят сложные процессы трансформации энергии, включающие миграцию энергии электронного возбуждения и транспорт электронов. В основе функционирования макромолекул лежит электронно-конформационные взаимодействия, которые проявляются в самых различных процессах, где участвуют биологически активные макромолекулярные комплексы. Тепловые движения атомов, входящие в полипептидную цепь, их повороты и вращения вокруг связей приводят к созданию большого количества внутренних степеней свободы. Это приводит к свертыванию цепи и образованию клубков макромолекул – макроскопические системы, в которых проявляются статический характер параметров (размеры, формы, степень свертывания). Как физический объект их своеобразие проявляются в сочетании как статических так и механических особенностей поведения макромолекул. С одной стороны большое число взаимодействий атомов создают большое количества степеней свободы и возможны создания различных конформаций, с другой стороны наличие химической связи между атомами ограничено -возможно образование конформеров. Взаимодействия атомов химической ковалентной связи определяет: 1. цепное строение биополимеров. 2. соединения друг с другом мономеров. Клетки и их органоиды – гетерогенные системы. Их существование и функционирование определяется межмолекулярными взаимодействиями нековалентного характера (взаимодействуют слабо, потому что сильные взаимодействия создали бы устойчивые жестские структуры, лишенные молекулярной подвижности, а молекулярная подвижность необходимо для выполнения различных задач (регуляция химических реакций, трансформация энергии). Слабое взаимодействие в биологических системах: 1) Вандервальсовы силы 2) Ионные связи 3) Водородные связи 4) Гидрофобные взаимодействия Первичная структура – линейная. Важную роль в конформации полипептидов играют вандевальсовые силы, гидрофбные взаимодействия, водородные связи. Вторичная структура. Пептидная цепь в белках имеет спиральную конфигурацию ( 1) Образование дисульфидных связей, которые могут соединить несколько спиралей между собой. В местах образования их ослабляется водородная связь и нарушется спирализация. 2) Наличие радикалов некоторых аминокислот, которые не укладываются в спираль и образуют отдельные складки, скрещивания водородных связей. Такие параллельно расположенные участки -
1) Белки с преобладанием 2) Белки, упакованы по типу 3) Смешанная вторичная структура Третичная структура – строго упорядоченная в пространстве укладка спирали и несколько участков цепи. Каждый белок имеет свою конфигурацию. Это связано с тем, что свободные карбоксильные, гидроксильные, аминные и другие группы боковых радикалов, взаимодействующих между собой с образованием амидных сложных эфирных связей. Водородные связи соединяют остатки двух соседних цепей и образуются дусульфидные мостики. Это делает структуру полужесткой. Четвертичная структура: Объединенные двух и более субъединиц. Третичная структура приводит к созданию сложной активной белковой молекулы. Гемоглобин: 4 глобулы Глобула формирует слабые взаимодействия (гидрофобные→ взаимодействия цепных молекул друг с другом). Собранные в пачки рассматриваются и образуются фибриллярные структуры, которые функционируют вне раствора и формируют надмолекулярные белковые структуры, которые состоят из большего количества макромолекул с невалентными взаимодействиями- это клеточные мембраны, хромосомы, глобулы, нити в молекулах. Нуклеиновые кислоты. Уотсон Крик – структура ДНК, которая представляет собой правовинтовую спираль, из двух полинуклеиновых цепей, при этом одна цепь обвита вокруг другой. Таким образом, пары гетероциклических оснований нах-ся внутри. Обе цепи соеденены между собой водородными связями, которые возникают между гетероциклическими основаниями.
|
|||||||
|
|
Последнее изменение этой страницы: 2017-02-19; просмотров: 463; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 13.58.25.81 (0.013 с.) |
 -спираль). Каждый атом H2 имеет избыточный положительный заряд, притягивающийся к отрицательно заряженному атому О2 в следущем витке спирали. Внутри образуются пептидные связи, а боковые радикалы аминокислот обращены наружу и могут взаимодействовать с молекулами окружающей среды. Спиральная конфигурация – вторичная. Структура полипептидной цепи спирализована неполностью. Инсулин – 60%. Причиной нарушения спирали являются:
-спираль). Каждый атом H2 имеет избыточный положительный заряд, притягивающийся к отрицательно заряженному атому О2 в следущем витке спирали. Внутри образуются пептидные связи, а боковые радикалы аминокислот обращены наружу и могут взаимодействовать с молекулами окружающей среды. Спиральная конфигурация – вторичная. Структура полипептидной цепи спирализована неполностью. Инсулин – 60%. Причиной нарушения спирали являются: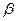 - структура,
- структура, 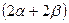 . В случае глобулы полипептидная цепь свернутая в клубок – третичная. Наличие нескольких сшивок S-S накладывает ограничение на конформацию.
. В случае глобулы полипептидная цепь свернутая в клубок – третичная. Наличие нескольких сшивок S-S накладывает ограничение на конформацию.


